| |
Chelodina longicollis: QM 59266-68, 59274, 59281-82, UC 0161, 0164, 0166. Chelodina oblonga: QM 59272-74, UC 0161-63. Chelodina expansa: QM 59284. Chelodina rugosa: QM 59264. Elseya dentata (Daly River, N.T.): NTM 13319, 13521, 16330, QM 59277-80, UC 0179. Elseya sp. aff. dentata (South Alligator River, N.T.): AM 128002, 128004, QM 59285-88. Elseya latisternum: AM 123037, 123039, 125474-75, QM 48054-55. Elseya sp. aff. latisternum (Manning River, N.S.W.): AM 123040, 123042, QM 59289-90. Elseya sp. aff. latisternum (Bellinger River, N.S.W.): AM 138387-88, UM 02016-17. Elseya novaeguineae (Sepik River, New Guinea): AM 42662, 125038. Emydura sp. aff. krefftii (Fraser Island, Qld.): QM 59275-76. Emydura sp. aff. subglobosa (Sleisbeck, N.T.): NTM 13428, 13433, UC 0171-72, 0177. Aspideretes hurum (no data): UC 0167. Chelus fimbriatus (Venezuela): PCHP 3985. Pelomedusa subrufa (no data): UC 0221. Phrynops gibbus (no data): UC 0222.
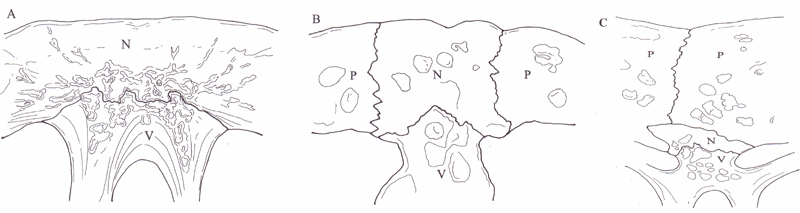
Well developed neural bones forming a contiguous series were observed in specimens of Aspideretes hurum (n=1, pleural pair VIII was in medial contact), Pelomedusa subrufa (n=1, pleural pair VIII was in medial contact), Chelus fimbriatus (n=1, pleural pair VIII was in medial contact), Phrynops gibbus (n=1, pleural pair I and V to VIII were in medial contact), Chelodina oblonga (n=6, pleural pairs I and VIII were in medial contact in all specimens with considerable variation for other pleural pairs. Table 1.) and Elseya sp. aff. latisternum (Manning) (n=4, pleural pairs I and VI to VIII were in contact in all specimens and pleural pair V in one specimen and II in two specimens) (Fig. 1). Neural bones were most developed in Chelus, being expanded both horizontally and vertically yielding much enlarged canals for the longissimus dorsi muscles. Rudimentary exposed neurals, small and isolated, were evident as individual variants in Chelodina longicollis (1 of 9, UC 0166), Elseya novaeguineae (1 of 2, AM 42662) and Elseya sp. aff. dentata (South Alligator River, N.T.) (1 of 6, QM 59285). Table 1 shows the neural formulae for all these specimens, following the conventions of Pritchard (1988).
No exposed neurals were evident in any of the specimens of Elseya latisternum (n=6), Emydura sp. aff. krefftii (n=2), Em. sp. aff. subglobosa (Sleisbeck) (n=5), Elseya sp. aff. latisternum (Bellinger) (n=4) (Fig. 1), Elseya dentata (n=8), Chelodina expansa (n=1) or Chelodina rugosa (n=1).
The presence of well developed exposed neurals in all four specimens of the undescribed species from the Manning drainage of New South Wales and their absence in all four individuals of its sister taxon (Georges and Adams, 1992) from the Bellinger River was a substantial and significant difference between these sibling taxa (Fisher Exact Test, p < 0.05).
A transverse section through the vertebral region of Asperedetes hurum revealed a suture between the neural bone and the underlying vertebral neural arch (Fig. 2A). Corresponding sections of a specimen of Chelodina longicollis with three exposed neurals revealed sutures similar to those observed in A. hurum (Fig. 2B), as did sections of C. longicollis, Elseya dentata and Em. sp. aff. subglobosa (Sleisbeck) (Fig. 2C) in the absence of exposed neural bones.
Discussion
This study establishes the undescribed Elseya from the Manning drainage of New South Wales as the second Australian chelid with well developed neural bones. The presence of neurals in this species and their absence in its sister taxon from the Bellinger River is a major discrete morphological difference in what was formerly a cryptic species pair (Georges and Adams, 1992). This species pair shows fixed differences at 20% of electrophoretic loci, despite little if any external morphological difference.
|
|
This provides an important example of where surveys based on molecular techniques can serve to focus attention on morphological features that might otherwise have gone undetected.
The Manning River Elseya has a rather short series of 3 to 5 neurals, a condition similar to that found in Phrynops gibbus (Chelidae). There is no obvious lateral expansion of the rib heads to accommodate enlarged longissimus dorsi muscles and in fact the ribs fit quite closely to the sides of the neural arches. This combination is also seen in Pelomedusa subrufa (Pelomedusidae) and is therefore considered to be the primitive condition.
If the function of well developed neurals is to add stability to shells particularly subject to lateral torsion (Pritchard, 1988) and this function is supplanted in chelids by the presence of a fixed pelvic girdle and extensive buttressing, then we would expect to see a correlation between the presence of neurals in chelid turtles and the lack of development of plastral buttresses. Indeed, anterior plastral buttressing is poorly developed in Chelodina oblonga, compared to other species of similar body form and habits in the Chelodina expansa group, and Chelodina oblonga has well developed neurals. Similarly, among the short-necked chelid turtles of Australia (excluding Pseudemydura), the Manning River form of Elseya which has well developed neurals, has the least developed anterior bridge buttresses.
On the basis of the bone sections, we suggest that there are three neural character states:
- Neural bones small, rudimentary, not visible in dorsal view being obscured entirely by the pleurals which meet medially for the full length of their common medial suture.
- Neural bones small, rudimentary, but exposed as small bony elements along the carapace midline. They do not form a contiguous series and pleural to pleural sutures make the predominant contribution to the midline suture.
- Neural bones well developed and dorsally exposed, forming a contiguous series of two or more. Pritchard (1988) has further subdivided this character state, based on a study of a greater range of specimens than examined here.
The demonstration of subsurface neural elements suturally separated from the neural arches of the dorsal vertebrae, with the possibility that neurals of some form may be present in all chelids, requires us to rethink our character definitions. The character state "neurals absent" should be instead "exposed neurals absent" and "neurals present" should become "neurals exposed". Also it will be necessary to appreciate that secondary development of exposed neurals may not imply reacquisition of a structure once lost but rather expansion of a persistent but rudimentary element.
Whether the subsurface neural elements are vestigial (that is, lacking function) is not clear. The possibility exists that by spanning the midline carapace suture from below, they reinforce it and relieve lateral pressure that would otherwise come to bear on the neural arches should the shell be subjected to downward force. Such a function would explain their retention.
Matching the distribution of well developed neurals among chelid turtles with current phylogenetic hypotheses is problematic (Gaffney, 1977). Neurals are well developed in the South American Chelus fimbriatus, Hydromedusa spp. and the Phrynops geoffroanus complex (Rhodin and Mittermeier, 1983; Pritchard, 1988), and in the Australian Chelodina oblonga (Burbidge et al. 1974) and Elseya sp. aff. latisternum (Manning) (present study). The character also shows great individual variability in Phrynops nasutus and P. gibbus (Pritchard, 1988). Clearly, either loss of exposed neurals has occurred independently many times, or well developed neurals have been secondarily derived independently many times, or a combination of the two is true.
|
|