| |
The most influential variable in the discrimination of females was HH (Partial R2 = 0.55; F=28.9, p<0.0001) followed by HWT (Partial R2 = 0.43, F=17.6, p<0.0001) and HL (Partial R2 = 0.34, F=11.2, p<0.0001), so differences in head shape were the most influential in providing overall discrimination between the three taxa. This is consistent with the fact that C. burrungandjii and the Kimberley form were first recognised as different species, by both Aboriginal people and Europeans, by differences in head shape. There was no clear partition of the raw variables in terms of their association with one or the other canonical variates, a situation not improved by varimax rotation, so we could not carry our interpretation further.
For males, the subset of variables that provided best discrimination were HWJ (Partial R2 = 0.53, F=12.2, p < 0.0005), HH (Partial R2 = 0.49, F=11.0, P<0.0005), VT (Partial R2=0.36, F=5.8, p<0.01) and HL (Partial R2 = 0.28, F=4.0, p<0.05). All are head measurements. A total of 92.5% of the among-groups variation was explained by the first canonical variate, which again provides the bulk of the discrimination between C. rugosa and the other two forms, but contributes little to the discrimination between C. burrungandjii and the Kimberley form (Figure 7b). In contrast to the analysis for the females, discrimination was not particularly good. Two of the 14 C. rugosa were misclassified, one each to C. burrungandjii and the Kimberley form. Two of the nine Kimberley males were misclassified as C. burrungandjii and two of the four male C. burrungandjii were misclassified as the Kimberley form. Hence, while the discrimination between C. rugosa and the other taxa is reasonably good, the distinction between C. burrungandjii and the Kimberley form breaks down for males. The distinction between these two may become more pronounced as they grow, and males grow to smaller sizes than females.
Distribution.—Chelodina burrungandjii is restricted to the Arnhem Land Plateau (Figure 8), a large sandstone plateau in the Northern Territory. It is found rarely in the plunge pools at the base of the Arnhem escarpment. Specimens have been collected from the upper reaches of the Mann, Liverpool, South Alligator, East Alligator and Katherine (Daly River drainage) and the Wilton rivers (Roper River drainage). The rivers of north-east Arnhem Land, that is, east of the Goyder River, contain Chelodina rugosa and Aboriginal people from the region do not appear to know of another form (B. Wununngmurra, pers. comm).
Etymology.—The specific epithet, burrungandjii, derives from the proper noun Burrungandji used by the Gagadju people of the western Arnhem Land region (Gundjeihmi language) to distinguish the turtle from all other turtles in the region (Lucas and Russel-Smith, 1993). We chose these names to recognise the long association between Aboriginal people and the turtle and their prior and detailed knowledge of the freshwater turtle fauna of Australia.
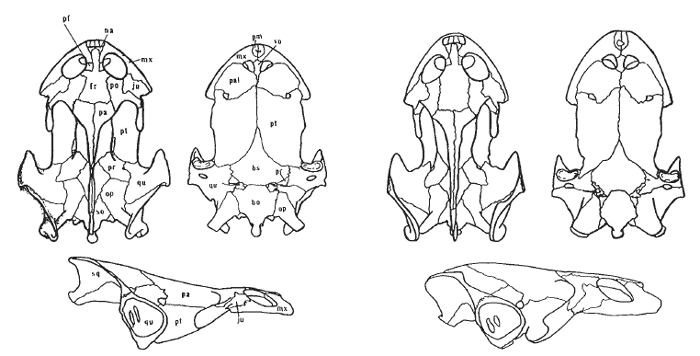
Related Taxa.—The nearest living relative of Chelodina burrungandjii is the undescribed Chelodina sp. (Kimberley). Their nearest relative is Chelodina rugosa, holotype: AM R6256 from Cape York, Queensland, Australia.
|
|
Description
External Morphology
Carapace.—The carapace (Figure 5) is oblong and flared over the hind legs in the region of M7-M9. There is a second smaller expansion of the shell at M5, although this is subject to ontogenetic variation. The widest point of the carapace is usually M8, and there is slight upturning of the marginals between M4 and M6. The scutes are smooth but there is a fine reticulated pattern throughout the carapace, and this is present in the underlying bone also. M2, M3 and M4 are the widest of the marginal scutes. A wide cervical scute is present. The first vertebral scute is significantly wider than the rest, a typical Chelodina condition. Vertebral formula is V1>V2>V3>V5>V4. In old specimens there is a marked medial furrow along the vertebral series. The V1/C1 sulcus contacts the middle of M3; C1/C2 sulcus contacts the posterior of M4; C2/C3 sulcus contacts the anterior of M7; C3/C4 sulcus contacts the anterior of M9 and the C4/V5 sulcus contacts M11. The carapace is dark brown to black, occasionally brown with extensive darker mottling and striations.
Plastron.—The plastron (Figure 6) is rectangular in general shape, in the sense that lines drawn to join the widest points of the anterior and posterior lobes of the plastron are roughly parallel. The widest point of the anterior lobe occurs toward its posterior extent, whereas that of the posterior lobe occurs in the middle. The scutes of the plastron are smooth, despite a fine reticulate pattern in the underlying bone. Plastral scute formula is: pect>int>fem>abd>hum>an>gul. The plastron and the ventral surfaces of the marginals are cream in colour with no darkening of scute margins. They are commonly stained caramel brown to orange in larger, slower-growing individuals.
Head and Soft Parts.—The dorsal to mid-lateral surface of the head is covered with smooth, soft skin broken into numerous irregular uncornified scales of negligible relief, dark olive green to black in colour, sometimes with fine black specks. Eyes are chocolate brown with gold, occasionally orange, ring bordering the pupil. Upper rhamphotheca olive with light black flecks. Lower rhamphotheca olive with numerous black/brown striations. Tympanum is light olive with a dark mottle. Barbels are variable in number, typically two are prominent, but up to four running in a line along the inside edge of each lower jaw. Ventral surface of head and throat in adults is covered with a loose granular skin. Granules beneath the head often line up to form striations radiating back from the apex of the chin. In most animals, the ventral surface of the head and neck is cream to white; in others, the granulations may be alternating grey and white to form a speckled pattern. The dorsal surface of the neck is dark olive green with a dark mottle, which occasionally coalesces to black and is covered in small low blunt tubercles. Dorsum of limbs and tail similar coloration to dorsum of neck; ventral surface of limbs and tail uniform cream with slight granulation.
|
|