| |
Crescent shaped scales on limbs coloured as per dorsum of limbs, even when extending onto the lighter latero-ventral surfaces. The inguinal and auxiliary pockets are cream to white, typically unremarkable; rarely with heavy grey/mauve blotches evenly spaced and extending back from the sides of the neck. In juveniles, the speckled pattern of the ventral surfaces of the head and neck is usually absent. Instead, the ventral surface of neck and limbs is cream as are the inguinal and axillary pockets.
Size and Sexual Dimorphism.—All laparoscoped males up to 185 mm were immature whereas all males of 197 mm and greater were mature, hence sexual maturity in males occurs at around 185 to 197 mm. All laparoscoped females up to 217 mm were immature whereas all females of 229 mm and greater were mature, hence sexual maturity in females occurs around 217 to 229 mm. The largest mature female recorded in this study was 271.3 mm and the largest mature male was 220.2 mm. The mean size of the five largest females was 250.9 + 2.3 mm and the mean size of the five largest males was 211.3 +2.8 mm, so the species shows sexual size dimorphism, as in other Chelodina. Mature males also have longer, thicker tails than females, a character that develops coincidentally with onset of sexual maturity, as in other chelid turtles (Georges, 1983; Kennett, 1996). . This species is smaller than C. rugosa, making it the smallest Australian member of the C. expansa group.
Osteology
Skull.—The skull of C. burrungandjii (Figs. 3-4) is highly flattened and shortened in comparison to other members of the C. expansa group (Table 3). The anterior head height of C. burrungandjii is approximately half that of the height of the tympanum whereas in C. rugosa these measurements are approximately equal. The skull is deeply emarginated both posteriorly and temporally to the extent that the parietal is almost absent. It exists only as an enlarged process on the dorsal surface of the squamosal. The processus trochlearis pterygoidei are visible in their entirety from above. The eye sockets are large, set dorso-laterally in the skull and are widely separated. Dorsally, the broadening of the frontal bone and the shortening of the maxillae (reducing the forward extent of the front of the skull) have yielded the shorter, wider skull shape. The inter-orbital length is similar to that of C. rugosa, however the diameter of the orbits is relatively smaller. Ventrally the palatines are shorter and broader than in C. rugosa and the vomer makes contact with the pterygoids in most specimens but divides the palatines in all specimens to some degree (Figure 3), a character that also distinguishes this species from C. expansa and the Chelodina sp. (Kimberley).
The basisphenoid is wider in comparison to other C. expansa group species and the quadrate is elongated from the medial contact with the basisphenoid to the lateral edge of the base of the tympanum. The quadrate is angled forward and is less robust than that of other species. The region of the squamosal into which the digastricus maxillae insert is large, and extends towards the braincase as is typical of C. expansa group species. Likewise, the region of the squamosal dorsal to the tympanic cavity is narrowed, to form a ridge, with no flattening or flaring. The crista paroccipitalis are narrow when viewed ventrally, a feature that is diagnostic for this species. The ventral process of the prefrontals is short in this species and contacts the maxilla but is restricted to the dorsal orbit.
Cervical Spine.—The central cervical articulation formula (sensu Williams, 1950) is the same as for all chelid turtles and this would appear to be a synapomorphy for the Chelidae: (2(, (3(, (4(, (5), )6), )7(, (8). The atlas-axis complex (sensu Hoffstetter and Gasc, 1969) comprises paired neural arches laterally, the first centrum ventrally and an intercentrum anteriorly. These units are completely fused into a single and elongated vertebra, and it is this unit which adds substantially to the neck length of the Chelodina.
|
|
The remaining cervicals are similar to each other in structure. The centra each possesses a sagittal blade, forming a series that is least developed in the eighth cervical and is largest in the second. The transverse processes are large, triangular in dorsal view, and occupy the posterior third of the centrum. Each postzygophosis is large and joins in the midline with its partner to give a semilunar shape to the zygophosis in cervicals three to eight.
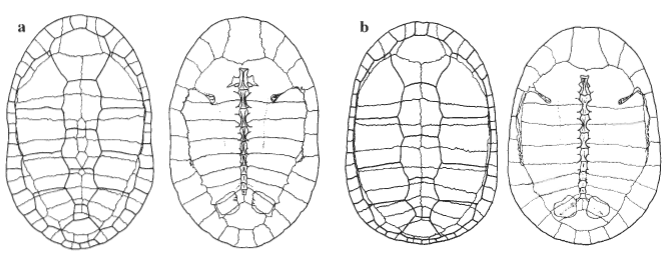
Shell.—The carapace (Fig. 5) is made up of the usual compliment of bones found in the Chelidae. The nuchal bone is approximately square in shape with elongation of marginals one to three and pleural pair one present, as is typical in the C. expansa group. Dorsally there is a usually a series of contiguous neural bones separating pleural pairs three to six. On examination of five shells neurals were always present, the number of exposed neurals ranged from three to six. The anterior bridge struts are enlarged and are at an angle of approximately 15° to the rib/gomphosis of pleural one. A medial expansion of the sutural surface is present with the rest of the suture equal in width to the marginal-plastral suture. Medially to the expansion of the anterior bridge strut is a large muscle attachment rugosity. This rugosity runs posteriorly to the midpoint of the fourth pleural and is the attachment point for the retrahens capitus collique muscles. The first four ribs are enlarged and expanded away from the vertebral column to make space for the enlarged longissimus dorsi muscles.
The gomphosis of pleural one inserts between peripherals four and five; pleural two inserts into the posterior third of peripheral five; pleural three into the posterior third of peripheral five; pleural four into the posterior third of peripheral six; pleural five into the posterior of peripheral seven; pleural six into the posterior of peripheral eight; pleural seven into the middle of peripheral nine; pleural eight into the middle of peripheral ten. Peripheral seven also contains the posterior bridge strut that does not make any contact with the pleurals.
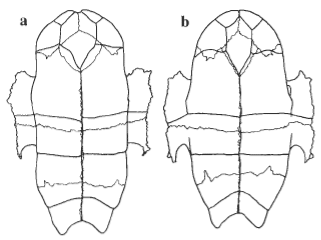
The plastron is unusual among Chelodina in that the intergular is extremely wide. The most anterior point of the plastron, at the junction of the epiplastra, is not ossified. This causes the complete separation of the gular scutes and the encroachment of the intergular on the plastral margin in many specimens.
 |
| |
Figure 6. Ventral views of the plastron of (a) Chelodina burrungandjii (UC2101) and (b) C. seibenrocki (UC0212). |
|
|
|